Why no one can say exactly how much is safe to drink while pregnant

I was waiting in the dining car of an Amtrak train recently when I looked up and saw that old familiar sign:
“According to the Surgeon General, women should not drink alcoholic beverages during pregnancy because of the risk of birth defects.”
One finds this warning everywhere: printed on bottles and menus or posted on placards at restaurants and even train cars barreling through Midwestern farmland in the middle of the night. The warnings are, of course, intended to reduce the number of cases of fetal alcohol syndrome in the United States. To that end, the Centers for Disease Control and Prevention (CDC) and the American Congress of Obstetricians and Gynecologists (ACOG) recommend that women avoid drinking any alcohol throughout their pregnancies.
Here’s how the CDC puts it:
“There is no known safe amount of alcohol to drink while pregnant.”
And here’s ACOG’s statement in 2008:
“. . . ACOG reiterates its long-standing position that no amount of alcohol consumption can be considered safe during pregnancy.”
Did you notice what they did there? These statements don’t actually say that no amount of alcohol is safe during pregnancy. They say that no safe amount is known and that no amount can be considered safe, respectively. Ultimately, these are statements of uncertainty. We don’t know how much is safe to drink, so it’s best if you don’t drink any at all.
Lest you think this is a merely a reflection of America’s puritanical roots, check out the recommendations of the U.K.’s National Health Service. While they make allowances for the fact that some women choose to drink, they still advise pregnant women to avoid alcohol altogether. As they say:
“If women want to avoid all possible alcohol-related risks, they should not drink alcohol during pregnancy because the evidence on this is limited.”
Yet it seems odd that the evidence is so limited. The damaging effects of binge drinking on fetal development were known in the 18th century and the first modern description of fetal alcohol syndrome was published in a French medical journal nearly 50 years ago. Six years later, in 1973, a group of researchers at the University of Washington documented the syndrome in The Lancet. Even then, people knew the cause of fetal alcohol syndrome: alcohol. And in the forty years since, fetal alcohol syndrome has become a well-known and well-studied illness. NIH alone devotes more than $30 million dollars annually to research in the field. So how come no one has answered the most pressing question (at least for pregnant women): How much is safe to drink?
One reason is that fetal alcohol syndrome isn’t like HIV. You can’t diagnose it with a blood test. Doctors rely on a characteristic pattern of facial abnormalities, growth delays and neural or mental problems – often in addition to evidence of prenatal alcohol exposure – to diagnose a child. Yet children exposed to and affected by alcohol during fetal development don’t always show all of these symptoms. Doctors and agencies now define fetal alcohol syndrome as the extreme end of a spectrum of disorders caused by prenatal alcohol exposure. The full spectrum, called fetal alcohol spectrum disorders (FASD), includes milder forms of the illness that involve subtler cognitive or behavioral problems and lack the classic facial features of the full-blown syndrome.
As you might imagine, milder cases of FASD are hard to identify. Pediatricians can miss the signs altogether. And there’s a fundamental difficulty in diagnosing the mildest cases of FASD. To put it crudely, if your child is slow, who’s to say whether the culprit is a little wine during pregnancy, genetics, too much television, too few vegetables, or god-knows-what-else? Unfortunately, identifying and understanding the mildest cases is crucial. These are the cases that worry pregnant women who drink lightly. They lie at the heart of the uncertainty voiced by the CDC, ACOG, and others. Most pregnant women would like to enjoy the occasional merlot or Sam Adams, but not if they thought it would rob their children of IQ points or otherwise limit their abilities – even just a little – down the line.
While it’s hard to pin down the subtlest cases in the clinic, scientists can still detect them by looking for differences between groups of children with different exposures. The most obvious way of testing this would be to randomly assign pregnant women to drink alcohol at different doses, but of course that experiment would be unethical and should never be done. Instead, researchers capitalize on the variability in how much women choose to drink during pregnancy (or at least how much they report that they drank, which may not always be the same thing.) In addition to interviewing moms about their drinking habits, the scientists test their children at different ages and look for correlations between prenatal alcohol exposure and test performance.
While essential, these studies can be messy and hard to interpret. When researchers do find correlations between moderate prenatal alcohol exposure and poor test performance, they can’t definitively claim that the former caused the latter (although it’s suggestive). A mysterious third variable (say, maternal cocaine use) might be responsible for them both. On the flip side, interpreting studies that don’t find correlations is even trickier. It’s hard to show that one thing doesn’t affect another, particularly when you are interested in very small effects. To establish this with any confidence, scientists must show that it holds with large numbers of people and that they are using the right outcome measure (e.g., IQ score). FASD impairments can span language, movement, math skills, goal-directed behaviors, and social interactions. Any number of measures from wildly different tests might be relevant. If a given study doesn’t find a correlation between prenatal alcohol exposure and outcome measure, it might be because the study didn’t test enough children or didn’t choose the right test to pick up the subtle differences between groups.
When studies in humans get tricky, scientists often turn to animal models. FASD research has been no exception. These animal studies have helped us understand the physiological and biochemical mechanisms behind fetal alcohol syndrome, but they can’t tell us how much alcohol a pregnant woman can safely drink. Alcohol metabolism rates vary quite a bit between species. The sensitivity of developing neurons to alcohol may differ too. One study used computational modeling to predict that the blood alcohol level of a pregnant rat must be 10 times that of a pregnant human to wreak the same neural havoc on the fetus. Yet computational models are far from foolproof. Scientists simply don’t know precisely how a dose in a rat, monkey, or other animal would translate to a human mother and fetus.
And here’s the clincher: alcohol’s prenatal effects also differ between humans. Thanks to genetic differences, people metabolize alcohol at very different rates. The faster a pregnant woman clears alcohol from her system, the lower the exposure to her fetus. Other factors make a difference, too. Prenatal alcohol exposure seems to take a heavier toll on the fetuses of older mothers. The same goes for poor mothers, probably because of confounding factors like nutrition and stress. Taken together, these differences mean that if two pregnant women drink the same amount of alcohol at the same time, their fetuses might experience very different alcohol exposures and have very different outcomes. In short, there is no single limit to how much a pregnant woman can safely drink because every woman and every pregnancy is different.
As organizations like the CDC point out, the surest way to prevent FASD is to avoid alcohol entirely while pregnant. Ultimately, every expecting mother has to make her own decision about drinking based on her own understanding of the risk. She may hear strong opinions from friends, family, the blogosphere and conventional media. Lots of people will seem sure of many things and those are precisely the people that she should ignore.
When making any important decision, it’s best to know as much as you can – even when that means knowing how much remains unknown.
_____
Photo Credit: Uncalno Tekno on Flickr, used via Creative Commons license
Hurley TD, & Edenberg HJ (2012). Genes encoding enzymes involved in ethanol metabolism. Alcohol research : current reviews, 34 (3), 339-44 PMID: 23134050
Stoler JM, & Holmes LB (1999). Under-recognition of prenatal alcohol effects in infants of known alcohol abusing women. The Journal of Pediatrics, 135 (4), 430-6 PMID: 10518076

 Humans learn about objects by exploring them. I
Humans learn about objects by exploring them. I 
 While we have five wonderful senses, humans rely most on our sense of sight. The allocation of real estate in the brain reflects this hegemony; a far greater chunk of your cerebral cortex is dedicated to vision than to any other sense. So when you encounter people, objects, and animals in the world, you typically use visual information to tell your lover from a toothbrush from your cat. And while it would be reasonable to expect your brain to process all of these items in the same way, it does nothing of the sort. Instead, the visual cortex segregates and plays favorites.
While we have five wonderful senses, humans rely most on our sense of sight. The allocation of real estate in the brain reflects this hegemony; a far greater chunk of your cerebral cortex is dedicated to vision than to any other sense. So when you encounter people, objects, and animals in the world, you typically use visual information to tell your lover from a toothbrush from your cat. And while it would be reasonable to expect your brain to process all of these items in the same way, it does nothing of the sort. Instead, the visual cortex segregates and plays favorites.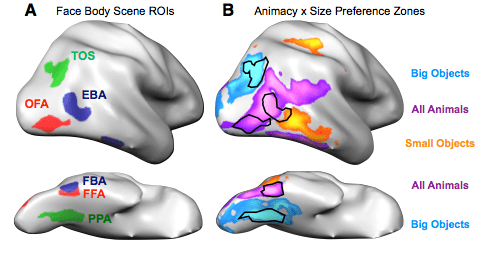
 More than a century ago, scientists discovered something usual about how people with
More than a century ago, scientists discovered something usual about how people with 