
Whenever I visit my childhood home outside of Chicago I try to make it to the local pancake house. The buttery pancakes would be reason enough, but they’re not the only reason I stop by. A stroll through that pancake house is truly a stroll down memory lane. Each table I pass triggers a memory of a meal shared with different people in different decades of my life. One moment I’m eating German pancakes with my college boyfriend. The next, I am passing menus to my new husband’s family. The next, I am celebrating my eighth grade graduation with my parents and older brother.
Memories return you to a specific time and place. Consider so-called flashbulb memories, or vivid memories of dramatic moments that caught you off-guard. I remember exactly where I was when I heard that a plane had struck one of the Twin Towers and, later, when I learned that my father had died. I remember that I was sitting on the living room rug in my Somerville apartment when I watched Columbia transform from a space shuttle into a streak of fire across the sky. Is it helpful to remember where I was sitting? Not in the slightest. But in the murky, mysterious realm of memory, when and what are inextricably linked with where.
Mention the word “memory” to neuroscientists and you’re sure to get them thinking of the hippocampus, a sliver of tissue nestled deep inside each hemisphere of the brain. The hippocampus has been synonymous with memory since the late 1950s, when William Scoville and Brenda Milner described a patient who was incapable of forming new memories after both of his hippocampi were removed. Since then, throngs of neuroscientists have devoted their careers to studying the hippocampus. Among other revelations, they’ve discovered a class of neurons called place cells that represent (you guessed it) information about place.
How do cells represent place? To illustrate, let’s say you’re in your favorite coffee shop. Some of the place cells in your hippocampus will fire like crazy when you walk through the entrance. Others save their enthusiasm until you are waiting in line to order your latte, stopping at the counter for milk and sugar, or settling in at your favorite table. When you physically occupy their place-of-interest, they go nuts – like a neural alarm signaling your location. At this moment, you are here!
The same principle applies to my experience at the pancake house. Different place cells fire at different tables. In essence, these sets of cells provide a unique neural code for each space I can occupy in the restaurant. And this code has been with me for a while. When I sat in the corner booth after my graduation from middle school, I formed a memory of that celebration that included the code for that particular spot. Decades later, sitting in that booth or even walking past it can trigger a similar code in my brain, one that elicits the rest of that dusty old memory.
While eternally cool, place cells have become old news in hippocampal research. The new hippocampal hotness is studying “time cells”. These recently discovered neurons prefer to fire at different intervals after an event (say, ten seconds versus one minute after you step into the coffee shop). This research fad is a bit amusing, as it turns out that place cells and “time cells” are one and the same. This fact hasn’t stopped scientists from referring to “time cells,” but it has forced them to typically use the term in quotation marks.
As scientists studied the time code in the hippocampal cells of rats, a flaw in their experiments became clear. Their studies recorded the neural activity of moving rats, which means that the firing patterns observed by the scientists could reflect changes in time, changes in the rat’s location, or in its motion.
Two recent papers addressed this issue and clarified the nature of “time cells” in the hippocampus. The first of these appeared in the journal Neuron in June of this year. The paper, by Benjamin Kraus, Michael Hasselmo, and collaborators at Boston University, describes an experiment that has as much to do with your time spent sweating it out at the gym as it does with your memory of past events. The scientists recorded the activity of hippocampal cells in rats as they ran on a treadmill or moved around in a simple maze. Since the rat remained in the same location as it ran on the treadmill, the researchers could decouple the rat’s location from the passage of time and the distance the rat ran. Since the authors could vary the speed of the treadmill, they could also piece apart the related variables of time and distance.
The scientists found that “time cells” still produced a time code when location was kept constant (on the treadmill). Using some fancy modeling, they also showed that the activity of most “time cells” reflected a combination of elapsed time and distance run, but a smaller number of “time cells” seemed to care only about time or distance. They also found that these same cells behaved like normal place cells when the rat walked around a simple maze. In short, place cells (a.k.a. “time cells”) can convey information about place, time, and distance travelled to varying degrees that also change under different conditions.
A second paper on the subject came out in a September issue of The Journal of Neuroscience. The authors, Christopher MacDonald, Howard Eichenbaum*, and colleagues (also from Boston University) eliminated the variable of location by physically restraining the rats from moving with a special headpiece that attached to the rats’ heads. This headpiece locked into the testing apparatus so that the rats couldn’t move their heads during testing. Unlike the fitness buff rats in the prior study, these rats were given a memory task. They got a whiff of an odor and then another whiff of an odor a few seconds later. If the second odor was the same as the first, the rat licked its waterspout and got a reward (a drop of water). If the two odors were different, the rat was not supposed to lick.
Even though the rats were completely immobile, the rats’ “time cells” showed a strong time code. Different cells fired at different times during the delay. These cells also seemed to represent what information (in this case, the odors presented for the task). The scientists found that the overall pattern of “time cell” firing was more similar when the rats remembered the same odor than when they remembered different odors across trials.
In short, place/time cells can represent what, when, and where in a variety of ways, depending on a variety of factors. This representation is flexible – just as memory must be in order for you to remember the date of your anniversary, the feel of your first kiss, and the items on your next shopping list. The remarkable thing about memory is that it is both flexible and robust, meaning that it is resistant to degradation or being swamped out by noise. It can return us to times, places, and experiences that are far away and decades past. For that, we can thank the hippocampus, neural codes, and a set of remarkable cells with an identity crisis.
_____
Photo credit: Stu Rapley on Flickr, used via Creative Commons License
*Howard Eichenbaum was also a middle author on the Neuron paper. Much of the recent work on “time cells” has come from his lab and affiliated labs at Boston University.
Kraus BJ, Robinson RJ 2nd, White JA, Eichenbaum H, & Hasselmo ME (2013). Hippocampal “time cells”: time versus path integration. Neuron, 78 (6), 1090-1101 PMID: 23707613
MacDonald CJ, Carrow S, Place R, & Eichenbaum H (2013). Distinct hippocampal time cell sequences represent odor memories in immobilized rats. The Journal of Neuroscience : the official journal of the Society for Neuroscience, 33 (36), 14607-14616 PMID: 24005311
 Humans learn about objects by exploring them. I
Humans learn about objects by exploring them. I 
 While we have five wonderful senses, humans rely most on our sense of sight. The allocation of real estate in the brain reflects this hegemony; a far greater chunk of your cerebral cortex is dedicated to vision than to any other sense. So when you encounter people, objects, and animals in the world, you typically use visual information to tell your lover from a toothbrush from your cat. And while it would be reasonable to expect your brain to process all of these items in the same way, it does nothing of the sort. Instead, the visual cortex segregates and plays favorites.
While we have five wonderful senses, humans rely most on our sense of sight. The allocation of real estate in the brain reflects this hegemony; a far greater chunk of your cerebral cortex is dedicated to vision than to any other sense. So when you encounter people, objects, and animals in the world, you typically use visual information to tell your lover from a toothbrush from your cat. And while it would be reasonable to expect your brain to process all of these items in the same way, it does nothing of the sort. Instead, the visual cortex segregates and plays favorites.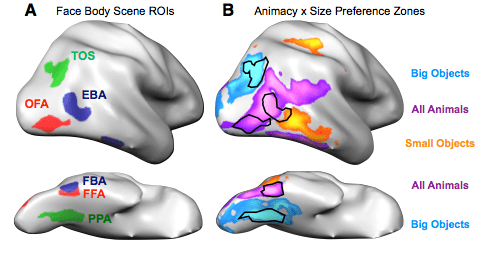
 More than a century ago, scientists discovered something usual about how people with
More than a century ago, scientists discovered something usual about how people with  Last month,
Last month, 